
|
Teaching |
|
I had lived in the same coastal town for around a year when I heard talk about strange comings and goings at a large cave on the beach. The beach is a 2 mile stretch of golden sand, with various caves. Legend has it that in this one particular cave things appear in the dead of night. I was keen to find out more, since all my life I have enjoyed exploring the great outdoors. I wanted to solve this mystery that was worrying the local population. One warm august evening, I managed to wait until the tide was out and walked down to the beach. I took my large lantern torch with me. I entered the cave through a small gap in the rock, only just wide enough for my rather large body. I switched on my Nitecore flashlight to find that I was surrounded by a large open space. My torch lit up a fine room moulded over millions of years by the sea. There were stalic tights and stalic mites everywhere. My lantern lit up the room and it looked like a fine cathedral. I walked around and found the most incredible colony of bats. There were hundreds of the fine, protected species, hanging splendidly from the ceiling. Some were flying around the room, and some more arrived just as I decided to leave. It really was an awesome sight, a real display of nature at its best. I could really understand why bats are such an important species in our ecosystem, just by looking at them from below. Rather than scare them with my big light to much I decided it was time to leave. Getting out of the cave was a little more difficult than getting in - I only just managed it! Next time I heard the discussions about what might be going on in the bat cave I decided to keep absolutely quiet. I think it was the right thing to do, to make sure this wonderful part of nature was not disturbed. |

|
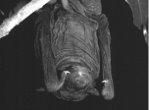
Hipposideros diadema |

|
Dr Tigga Kingston |
|
Assistant Professor |


|
Cheiromeles torquatus |